Fibroblasts are widely distributed stromal cells in the body, with core functions including maintaining tissue architecture, regulating extracellular matrix (ECM) homeostasis, and participating in tissue damage repair and inflammatory responses. During tumor development, normal fibroblasts are activated by abnormal signaling from the tumor microenvironment, transforming into cancer-associated fibroblasts (CAFs), participating in the construction of a disordered tumor wound repair microenvironment, and serving as key cellular components that shape the tumor microenvironment and drive tumor progression.
Early studies simply classified CAFs into two types: inflammatory CAFs (iCAFs) and myofibroblastic CAFs (myCAFs), with understanding limited to their basic phenotypes and single functions. With advances in single-cell transcriptomics, ultra-high-throughput spatial sequencing, and lineage tracing technologies, the scientific community has confirmed that CAFs do not have a unified, stable phenotypic characteristic but possess extreme heterogeneity and plasticity, which is also the core reason for the failure and poor efficacy of traditional broad-spectrum CAF-targeted therapies.
The heterogeneity of CAFs is not driven by genetic variations such as gene mutations or chromosomal abnormalities, but relies on transcriptional reprogramming and epigenetic remodeling, dynamically regulated by multiple factors including tumor cell mutation status, microenvironment signals, spatial localization, and host status. Their plasticity manifests as different CAF subpopulations undergoing phenotypic and functional interconversion with tumor progression and environmental changes, widely participating in the entire process of tumor immune evasion, stromal remodeling, angiogenesis, neural infiltration, and treatment resistance. Therefore, systematically analyzing the layered characteristics of CAF heterogeneity, the regulatory mechanisms of plasticity, and clarifying their role in tumor therapy is of significant academic value and clinical importance for developing precise stromal-targeted anti-tumor strategies and breaking through the bottlenecks of solid tumor treatment.
There is significant spatial structural heterogeneity within solid tumors, with obvious differences in cellular composition, oxygen concentration, nutrient supply, mechanical tension, inflammation levels, and immune cell infiltration in different anatomical regions, forming functionally specific tumor ecological niches. The microenvironment signal characteristics of different niches vary significantly, which can directionally induce fibroblasts to differentiate into functionally specific CAF subtypes, ultimately forming location-specific CAF subpopulations. CAFs in each niche interact with surrounding cells through unique ligand-receptor signaling networks, exerting completely different biological functions, directly affecting local tumor progression, immune status, and patient clinical prognosis, serving as an important spatial basis for CAF functional heterogeneity.
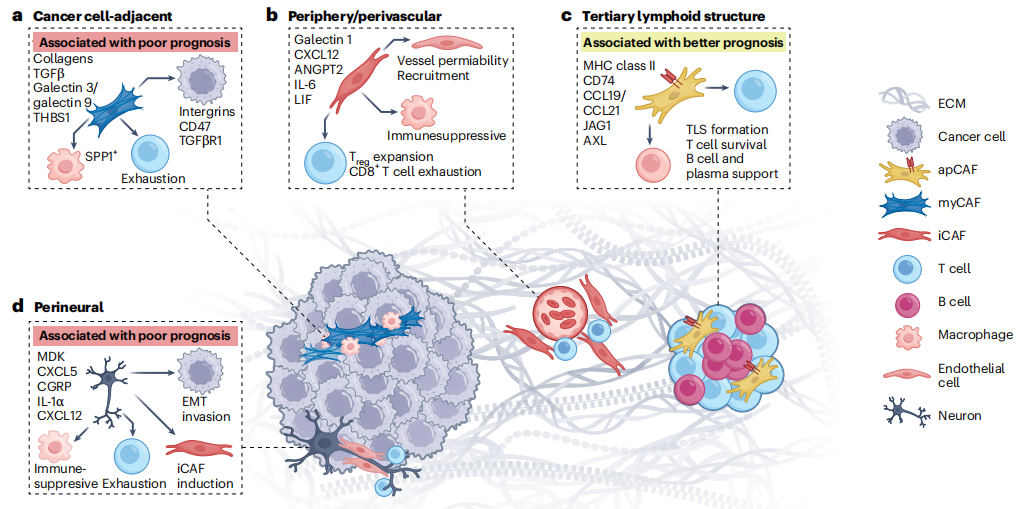
Cross-cancer integrated analysis has identified four conserved CAF characteristic niches and corresponding cellular subpopulations:
1. Cancer Cell Proximity Niche: This region has myCAFs as the absolute dominant subtype. These CAFs highly express type I/III/IV collagen, core TGFβ pathway components (TGFβ, THBS1), and matrix metalloproteinases (MMP1, MMP11) and other functional molecules. The core biological function is to massively remodel ECM, significantly increase tumor tissue stiffness, and promote tumor cell epithelial-mesenchymal transition (EMT) and local invasion. Meanwhile, this subtype can induce SPP1+ M2-type macrophage polarization, CD8+ T cell exhaustion and functional inhibition in the microenvironment by secreting key factors such as Galectin-3, Galectin-9, CXCL12, and TGFβ, constructing a typical immune-excluded, immunosuppressive microenvironment. Extensive clinical sample validation shows that high enrichment of myCAFs in this niche is closely related to poor tumor prognosis and primary resistance to immune checkpoint inhibitors.
2. Tumor Peripheral/Perivascular Niche: Tumor invasive margins and areas around blood vessels are mainly enriched with iCAFs subtype. These cells highly express complement system components, inflammatory cytokines, and chemokines, including core molecules such as IL-6, IL-11, and C3. iCAFs are closely adjacent to vascular endothelial cells and myeloid immune cells, and can regulate the tumor microenvironment through multiple pathways: on one hand, secreting factors such as VEGFA and FGF2 to promote tumor angiogenesis and maintain vascular stability; on the other hand, recruiting regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs) through signals such as IL-6 and CXCL12, inhibiting the anti-tumor function of cytotoxic T cells and mediating immunosuppression. Notably, the impact of iCAFs on patient prognosis has signal dependency and tumor specificity. In some tumors, they can mediate poor prognosis and drug resistance, while in some cancers, they may also exhibit moderate anti-inflammatory and homeostasis-regulating protective effects, showing significant functional duality.
3. Tertiary Lymphoid Structures (TLSs) Niche: Tertiary lymphoid structures are important functional immune micro-regions within tumors, with this niche specifically enriched with antigen-presenting CAFs (apCAFs). Limited by signal overlap interference from traditional multiplex immunofluorescence techniques, apCAFs have been difficult to precisely identify for a long time. Recent advances in lineage tracing and single-cell spatial multi-omics technologies have successfully achieved precise localization and functional analysis of this subtype. apCAFs highly express key molecules such as MHC II, CD74, CCL19, and CCL21, can directly complete antigen processing and presentation, activate CD4+ helper T cells, recruit T and B lymphocytes, maintain plasma cell homeostasis, and promote TLS maturation and adaptive anti-tumor immune activation. Clinical data confirm that patients with high apCAF enrichment have better prognosis and significantly improved response rates to immune checkpoint inhibitor therapy, making them key tumor-suppressive stromal cells in the tumor microenvironment.
4. Perineural Niche: Nerve-tumor microenvironment interaction is an emerging hot topic in tumor research in recent years and an important regulatory pathway for malignant progression of solid tumors. The perineural niche is mainly composed of tumor-associated nerves, Schwann cells, CAFs, and tumor cells, forming a complex signal interaction network. Tumor cells can actively reprogram nerve cells, promoting tumor neural infiltration and new nerve fiber generation; activated Schwann cells and nerve cells can secrete factors such as IL-1α, CXCL5, and CXCL12, inducing local CAFs to transform toward a pro-tumor phenotype. These CAFs can further promote tumor EMT progression, enhance cell invasion and migration capabilities, induce macrophage M2 polarization, and inhibit local anti-tumor immunity, ultimately forming a nerve-dependent pro-tumor microenvironment, highly correlated with high tumor invasiveness, recurrence, and poor prognosis. Currently, this field is still dominated by mouse model studies, and mechanistic verification in human clinical samples remains to be further improved.
This review integrates multi-cancer single-cell and spatial multi-omics data, mouse lineage tracing models, and functional intervention experimental results, systematically constructing a CAF heterogeneity research system from three mutually coupled, progressively deepening dimensions: transcriptomic phenotypes, cellular origin lineages, and molecular marker expression. Research has confirmed that CAF subtype differentiation, phenotypic characteristics, and functional output are not determined by a single factor, but are the result of combined effects of spatial localization, innate lineage, transcriptional regulation, and microenvironmental selection. Heterogeneity at different dimensions mutually superimposes and regulates each other, ultimately shaping a highly complex, dynamically variable tumor stromal cell ecosystem, serving as an important stromal basis for individual differences in tumor progression and treatment response.
Tumor tissues have significant spatial heterogeneity, with obvious differences in cell composition, nutrient supply, mechanical stress, and immune infiltration in different regions, forming specific tumor ecological niches that shape location-specific CAF subpopulations. CAFs in each niche have completely different signal interactions, functional effects, and clinical prognosis, serving as the spatial basis for CAF functional differentiation.
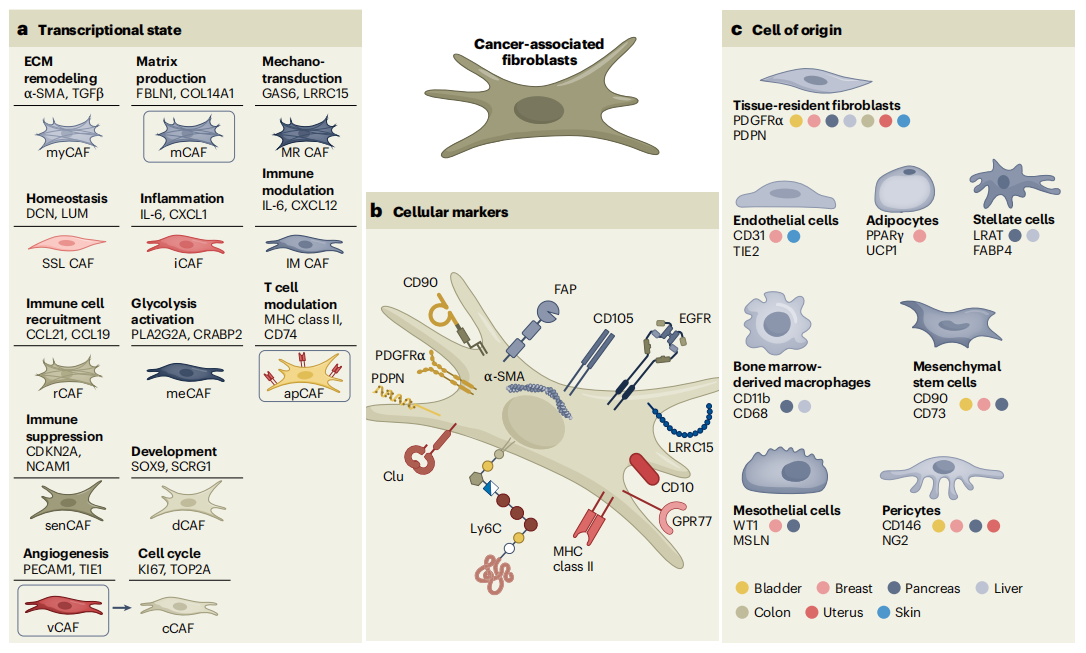
● Classical core subtypes: myCAFs are centered on ECM remodeling, relying on TGFβ signaling pathway activation; iCAFs are centered on inflammatory immune regulation, driven by IL-1/JAK/STAT signaling, and the two can dynamically convert with microenvironment signals. apCAFs highly express MHC II molecules and antigen presentation-related genes, mainly participating in immune activation, distinguished from classical pro-tumor subtypes.
● Novel functional subtypes: Studies have successively identified special subpopulations such as vascular CAFs (vCAFs), metabolic CAFs (meCAFs), senescent CAFs (senCAFs), and reticular CAFs (rCAFs). Among them, vCAFs regulate tumor angiogenesis, meCAFs mediate immunosuppression through metabolic reprogramming, senCAFs promote tumor progression through senescence-associated secretory phenotype, with each subtype having independent transcriptional characteristics and non-overlapping functions, enriching the CAF heterogeneity system.
Based on single-cell transcriptomic sequencing data, this review breaks through the traditional iCAFs/myCAFs binary classification system, identifying multiple CAF subpopulations with specific transcriptional characteristics and functions, confirming that CAF transcriptional phenotypes have high tissue conservation and tumor specificity.
● Tissue-resident fibroblasts: These are the most important origin cells of CAFs. Resident fibroblasts from different tissues and subtypes are activated by tumor paracrine signaling stimulation, forming functionally different CAFs. For example, pancreatic stellate cells and peribiliary fibroblasts can both differentiate into CAFs, but with completely different transcriptional phenotypes.
● Vascular wall stromal cells: Pericytes and other vascular wall stromal cells can transdifferentiate into CAFs, divided into two types: fibrotic pericytes and perivascular cells, mediating ECM remodeling and angiogenesis respectively, serving as an important source of CAFs in the tumor vascular microenvironment.
● Mesothelial cells: Mainly originating from body cavity mesothelial cells, they are important precursor cells of apCAFs, enriched in pancreatic cancer, ovarian cancer, and peritoneal metastatic cancer, specifically regulating the tumor immune microenvironment.
● Other origin cells: Bone marrow-derived cells, endothelial cells, adipocytes, and mesenchymal stem cells can all form CAFs through transdifferentiation and migration activation. These exogenous-origin CAFs often have stronger pro-metastatic and pro-drug-resistance capabilities.
CAF markers have no absolute universality; their expression dynamically changes with tumor type, progression stage, and microenvironmental status. The same marker can be differentially expressed in different subtypes, forming heterogeneity at the marker level, while also providing a basis for subtype identification and targeted screening.
● Pan-CAF markers: PDPN, PDGFRα, CD90, and others can broadly label most CAFs, but with tissue-specific differences. For example, PDPN only specifically labels iCAFs and myCAFs in breast cancer and squamous cell carcinoma, and cannot label apCAFs.
● Subtype-specific markers: α-SMA and LRRC15 specifically label myCAFs, participating in ECM remodeling and immunosuppression; IL-6 and CLU are characteristic markers of iCAFs, mediating inflammatory signaling and immune evasion; MHC II and CD74 are core markers of apCAFs, dominating antigen presentation functions; FAP can label various pro-tumor CAF subpopulations, participating in tumor metastasis regulation.
● Functional classification markers: CD105 can distinguish pro-tumor and tumor-suppressive CAFs. CD105+ CAFs promote tumor growth, while CD105- CAFs can activate adaptive immunity to inhibit tumor progression.
Static heterogeneity of CAFs is determined by cellular innate lineage characteristics, while dynamic plasticity completely depends on multi-level signal regulation from tumors and microenvironment, representing an important manifestation of tumor adaptive evolution. This review integrates multi-omics and functional experimental evidence to systematically classify the core driving mechanisms of CAF heterogeneity and plasticity into three major levels: local signal regulation of the primary tumor microenvironment, specific remodeling of distant metastatic microenvironment, and systemic regulation of host overall status. It comprehensively elucidates the molecular mechanisms of CAF phenotypic conversion and functional remodeling from local to overall, from static to dynamic, clarifying the multi-level regulatory network of CAF plasticity.
● Tumor cell genetic characteristics: The gene mutation subtypes and driver gene status of tumor cells are the upstream core factors regulating CAF differentiation direction, which can specifically shape stromal cell phenotypes through paracrine signaling. For example, tumors with BRCA gene mutations can specifically induce massive expansion of CLU+ iCAFs by activating stromal HSF1 signaling pathway, forming an inflammatory pro-tumor microenvironment; loss of TP53 function or mutation can induce CAFs to highly express proteoglycans and other stromal remodeling factors, promoting tumor invasion, metastasis, and chemotherapy resistance; pancreatic basal-like tumor cells can form bidirectional positive feedback regulatory loops with surrounding myCAFs, continuously enriching fibrotic and immunosuppressive pro-tumor microenvironment, accelerating malignant tumor progression.
● Microenvironment signal gradients: TGFβ and IL-1 signal gradients are classic core pathways regulating CAF bidirectional plasticity. The dynamic balance of signal intensity between the two directly determines CAF subtype differentiation. High concentration TGFβ signaling can significantly promote iCAF-to-myCAF conversion, enhancing stromal fibrosis and immune exclusion; while IL-1-mediated JAK/STAT inflammatory signaling activation can reverse this process, inducing myCAF-to-iCAF conversion and increasing microenvironmental inflammation levels. This signal gradient-dependent phenotypic conversion is the core mechanism for CAFs to adapt to local microenvironmental changes and achieve dynamic plasticity.
● Metabolic microenvironment regulation: The metabolic state of the tumor microenvironment significantly affects CAF phenotype and function. Hypoxia can induce CAFs to adopt a pro-tumor phenotype through HIF-1α pathway activation; nutrient deprivation can drive metabolic reprogramming of CAFs, shifting them toward a phenotype that supports tumor growth. Meanwhile, metabolic waste products such as lactic acid and reactive oxygen species in the microenvironment can also regulate CAF function and differentiation.
● ECM mechanical characteristics: The mechanical properties of the tumor stroma, including stiffness, architecture, and composition, are important factors regulating CAF phenotype. Increased ECM stiffness can activate mechanosensitive signaling pathways in CAFs (such as YAP/TAZ), promoting myCAF differentiation; while ECM degradation or softening may favor iCAF phenotype maintenance. This mechanical signal-dependent CAF phenotypic regulation provides new targets for understanding and intervening in tumor-stromal interactions.
CAFs at metastatic sites exhibit distinct phenotypic characteristics and functional adaptations compared to primary tumor sites, with significant differences in origin, phenotype, and function. Pre-metastatic niche formation is a key mechanism through which CAFs influence tumor metastasis.
● Organ-specific CAF subpopulations: CAFs at different metastatic sites show high organ specificity. For example, bone-metastatic CAFs highly express bone-related factors (such as CXCL12, SPP1), promoting osteoclast differentiation and bone destruction; liver-metastatic CAFs express metabolic regulators, creating a microenvironment conducive to metastatic tumor cell colonization; lung-metastatic CAFs express molecules related to vascular remodeling and immune regulation, promoting extravasation and survival of metastatic tumor cells.
● Recruitment and activation of exogenous CAFs: Tumor-derived exosomes and secreted factors can recruit bone marrow-derived cells and mesenchymal stem cells to distant sites, where they differentiate into CAFs with pro-metastatic characteristics. These exogenous CAFs often express higher levels of ECM remodeling factors and inflammatory cytokines, creating a microenvironment that supports tumor metastasis and colonization.
● Metabolic coupling between CAFs and metastatic tumor cells: At metastatic sites, CAFs often undergo metabolic reprogramming, shifting toward glycolysis or other metabolic pathways, providing metabolic support for metastatic tumor cells through the "reverse Warburg effect" or other metabolic coupling mechanisms, promoting metastatic tumor cell survival and proliferation.
Beyond local microenvironmental regulation, systemic host factors also significantly affect CAF heterogeneity and plasticity. These systemic factors integrate signals from the whole body, achieving broader regulation of tumor stromal cells.
● Aging-related changes: Aging is an important factor affecting CAF function and phenotype. Senescent CAFs (senCAFs) accumulate in the tumor microenvironment with age, characterized by senescence-associated secretory phenotype (SASP), secreting inflammatory cytokines, growth factors, and proteases that promote tumor progression. Meanwhile, aging-related changes in systemic hormone levels and immune function also indirectly affect CAF activity.
● Obesity and metabolic syndrome: Obesity is associated with changes in CAF phenotype and function. Adipose tissue-derived CAFs in obese individuals often exhibit enhanced pro-tumor activity, secreting more inflammatory factors and metabolic regulators. Additionally, systemic inflammation caused by obesity creates conditions favorable for CAF activation and tumor progression.
● Microbiome regulation: The gut microbiome and local tumor microbiome significantly affect CAF function through metabolites, bacterial components, and immune modulation. Specific bacterial species can induce CAF activation and pro-tumor phenotype through TLR signaling and other pathways, while microbiome-derived short-chain fatty acids and other metabolites can regulate CAF inflammatory response and immune modulation functions.
● Circadian rhythm and sleep: Emerging evidence suggests that circadian rhythm disruption and sleep disorders affect CAF function. Clock genes in CAFs regulate their metabolic status, inflammatory response, and interactions with tumor cells, potentially linking systemic lifestyle factors to tumor progression through CAF-mediated pathways.
Understanding the heterogeneity and plasticity of CAFs has important clinical implications for tumor diagnosis, prognosis assessment, and treatment selection. This review discusses the clinical applications and future therapeutic strategies targeting CAFs based on CAF heterogeneity.

The composition and spatial distribution of different CAF subpopulations have significant prognostic value. Specific CAF subtypes are closely associated with patient survival, treatment response, and drug resistance.
● myCAF enrichment is associated with poor prognosis: High myCAF content in tumor stroma is often associated with ECM remodeling, immune exclusion, and poor prognosis. This association is particularly evident in pancreatic cancer, breast cancer, and colorectal cancer.
● apCAF enrichment predicts better prognosis: Patients with high apCAF content show better response to immunotherapy and longer survival. The presence of apCAFs indicates an active anti-tumor immune microenvironment.
● CAF spatial distribution patterns: Beyond simple abundance, the spatial distribution of CAFs (such as location near cancer cells, proximity to blood vessels, or within TLSs) has additional prognostic value, providing more refined stratification for patient prognosis assessment.
Based on CAF heterogeneity, targeted therapeutic strategies need to be refined and personalized. The following are main therapeutic approaches:
● Promoting tumor-suppressive CAF conversion: Rather than simply eliminating all CAFs, the goal is to reprogram pro-tumor CAFs into tumor-suppressive phenotypes. TGFβ pathway inhibitors, vitamin A derivatives (retinoids), and other agents can induce myCAF-to-apCAF conversion.
● Targeting specific CAF subpopulations: Developing subpopulation-specific targeting strategies based on unique markers of different CAF subtypes. For example, FAP-targeted therapy can selectively eliminate FAP+ pro-tumor CAFs while preserving tumor-suppressive CAFs.
● Restoring anti-tumor immunity: Enhancing the antigen-presenting function of apCAFs or promoting TLS formation to improve anti-tumor immune responses. Combination therapy with immune checkpoint inhibitors may enhance treatment efficacy.
● Normalizing CAF-mediated ECM: Reducing excessive ECM deposition and tissue fibrosis to improve drug delivery and reduce immune exclusion. LOXL2 inhibitors, MMP inhibitors, and other agents can normalize the stromal barrier.
Despite significant advances in understanding CAF heterogeneity, several challenges remain:
● Lack of universal markers: Current CAF markers lack specificity and universality. Future research needs to identify more precise subpopulation-specific markers through single-cell multi-omics and spatial transcriptomics.
● Dynamic plasticity complexity: CAF phenotypic conversion is highly plastic and context-dependent. Understanding the dynamic regulatory mechanisms of CAF plasticity is crucial for developing effective intervention strategies.
● Clinical translation gaps: Many CAF-targeted therapies have shown efficacy in preclinical models but have not achieved expected results in clinical trials. This highlights the need for better translational models and more precise patient stratification.
● Combination therapy optimization: The optimal combination of CAF-targeted therapy with existing treatments (chemotherapy, immunotherapy, targeted therapy) remains to be determined. Future clinical trials need to explore rational combination strategies based on CAF heterogeneity.
This review comprehensively summarizes the latest advances in understanding CAF heterogeneity and plasticity. CAFs are not a uniform cell population but a highly heterogeneous and dynamically variable cell ecosystem, with their phenotypes and functions shaped by multiple factors including spatial localization, cellular origin, and microenvironmental signals.
The spatial distribution heterogeneity of CAFs in the tumor microenvironment provides an important structural basis for their functional diversity. The four characteristic niches identified (cancer cell proximity niche, tumor peripheral/perivascular niche, TLSs niche, and perineural niche) each host specific CAF subpopulations that exert distinct biological functions through unique signaling networks.
The core driving mechanisms of CAF heterogeneity and plasticity involve multi-level regulation from primary tumor microenvironment to distant metastatic microenvironment and systemic host status. Understanding these regulatory networks provides a theoretical basis for developing precise CAF-targeted therapeutic strategies.
Future research should focus on: (1) developing more precise CAF subpopulation identification and targeting technologies; (2) deeply exploring the dynamic plasticity mechanisms of CAFs; (3) optimizing CAF-targeted therapeutic strategies and combination approaches; (4) promoting clinical translation of CAF-related research findings. With advances in single-cell and spatial technologies and the accumulation of clinical evidence, CAF-targeted therapy is expected to become an important component of precision tumor therapy.
In summary, CAF heterogeneity and plasticity represent both challenges and opportunities for tumor therapy. By precisely understanding and intervening in CAF subpopulations, we can develop more effective anti-tumor strategies, bringing new hope for cancer patients.
References
Pramod, S., Lavon, H., Scherz-Shouval, R. et al. Heterogeneity and plasticity of cancer-associated fibroblasts. Nat Cancer 7, 583–596 (2026). https://doi.org/10.1038/s43018-026-01146-x